Hjem » Pulmonal patofysiologi » Transport af kuldioxid i blodet
Transport af kuldioxid i blodet
Måske fordi ilt er livets molekyle, er en betragtelig større andel af den gennemsnitlige fysiologistuderendes mentale kapacitet tilegnet iltens transport i blodet end kuldioxids transport i blodet. Måske fordi man dør hurtigere af hypoxisk respirationssvigt end af et hyperkapnisk svigt?
Det skal vi i hvert fald forsøge at gøre til skamme. For nok er ilt livets molekyle, men spørger man biologerne, så hedder det jo kulstofskredsløbet, og for hele verdens flora er kuldioxid jo inputtet og ilt det ekskretoriske output. Så for at square the circle, her følger om transport af kuldioxid i blodet.
Alle lærebøger i medicinsk fysiologi vil have enten et kapitel eller et længere afsnit tilegnet spørgsmålet om kuldioxids transport i blodet. Det følgende læner sig op af Wests klassiske The Essentials of Respiratory Physiology: Mest fordi en af de mest centrale figurer i nedenstående er planket fra Boron & Boulpaep, der selv har planket den fra West, men som også genfindes i Søren Lyagers Respirationsfysiologi og sikkert også andre steder.
Indhold i det følgende er:
- Kuldioxids forskellige former i Volumen og mængde af transporteret kuldioxid
- Volumen og mængde af transporteret kuldioxid
- Forskelle mellem CO2 i arterielt og i venøst blod
- Haldane-effekten og carbamino-forbindelser
- Carboanhydrase og CO2
- CO2-dissociationskurven
- Kilder
Kuldioxid findes på tre (fem) forskellige former
Det vil fremgå klart af både tabeller og figurer nedenfor, at CO2 findes på tre forskellige former i blodet. Det er ikke helt rigtigt. Der findes mikromolære koncentrationer af kulsyre og karbonat, som jeg i det følgende tillader mig at ignorere:
- CO2 findes som som fri, opløst gas i en mængde på godt 1 mM, et volumen på 2,4 mL/dL. Det udgør 5 % af den totale CO2 i kroppen. Det er denne frie, opløste gas, der er årsag til partialtrykket af CO2 i blodet på omtrentligt 40-46 mmHg
- CO2 findes som bikarbonat i en mængde på 24 mM, et volumen på 43 mL/dL. Det udgør 90 % af den totale CO2 i kroppen. Det er bikarbonat, der driver værket! Det er blodets ækvivalent til den internationale skibstrafiks 20 fodscontainer!
- CO2 findes bundet til hæmoglobin som karbamino-forbindelser. Det er godt 1 mM eller 2,4 mL/dL og igen omtrentligt 5 % af kroppens totale CO2. Karbamino-forbindelser, you might ask, det er de der uspecifikke amin-grupper på proteiner, der kan snuppe et kuldioxid. Det er baggrunden for Haldane-effekten, hvorom der skal pippes mere nedenfor.
Volumen og mængde af transporteret kuldioxid
Det er værd at dvæle lidt ved den faktiske mængde og volumen af kuldioxid, vi taler om, når vi taler om kuldioxids transport. Her er Boron & Boulpaeps eTabel 29-1 i kapitel 29 om transport af gasser i blodet uforlignelig, som det lønner sig at gengive noget af indholdet af i en tabel herunder.
Ved fysiologisk tryk og temperaturforhold giver det et volumen på 48 mL CO2 per dL arterielt blod og 52 mL CO2 pr dL venøst blod.
| Arterielt blod | Blandet-venøst blod | |||||
|---|---|---|---|---|---|---|
| Konc (mM) | Bidrag til total CO₂ (mL/dL) | Andel af total CO₂ (%) | Konc (mM) | Bidrag til total CO₂ (mL/dL) | Andel af total CO₂ (%) | |
| Fysisk opløst kuldioxid (CO₂) | 1,2 | 2,4 | 5 | 1,4 | 2,8 | 5,3 |
| Bikarbonat (HCO₃⁻) | 24 | 43,2 | 90 | 25,6 | 46,0 | 88,5 |
| Karbamino-forbindelser | 1,2 | 2,4 | 5 | 1,6 | 3,2 | 6,2 |
| Total | 26,4 | 48 | 100 | 28,6 | 52 | 100 |
Der er omtrentligt 26,4 mmol CO2 per liter arterielt blod og 28,6 mmol CO2 per liter blandet venøst blod. Ved fysiologiske tryk og temperaturer svarer det til, at der er opløst et volumen på omtrentligt 48 mL CO2 per dL arterielt blod og 52 mL CO2 per dL venøst blod. Man ser altså, at det totale indhold af CO2 varierer med arterielt og venøst blod, sådan at koncentrationen af CO2 i venøst blod er omtrentligt 2,2 mM højere end i arterielt blod. Det skyldes selvfølgelig, at metabolismens slutprodukt er CO2.
Hvordan når man fra 26,4 mM til 48 mL/dL, kunne man finde på at spørge?
Den opløste CO2, de 1,2 mM, kan selvfølgelig omregnes til et volumen ved hjælp af idealgasligningen, men hvordan skal man omregne bikarbonat og carbamat – der ikke er gasser – til et volumen? I helt gamle dage (Van Slyke & Neill i 1924) tog man blod, overhældte det med mælkesyre, sådan at al bikarbonat og alle karbaminoforbindelser blev omdannet til CO2. Med andre ord tog man følgende ligevægt:
CO₂ + H₂O ↔ H₂CO₃ ↔ H⁺ + HCO₃⁻
Og forskød den helt og aldeles mod venstre. Herefter brugte man en trykmåler til at måle CO2-trykket af en kendt volumen blod, sådan at man kunne estimere den molare stofmængde med idealgasligningen. Der er selvfølgelig flere detaljer til historien, men lidt har også ret.
Take home-message: Fra en uvægtet gennemsnitsbetragtning er der 27,5 mmol CO2 per liter blod og 50 mL CO2 per dL blod. I et normalfysiologisk individ (du ved, dem med 5 L blod i kroppen) giver det 137,5 mmol CO2 og 2,5 L CO2.
Så længe man ikke er død, producerer kroppen evindeligt mere CO2, og det skal cleares. Forudsætter man en normalfysiologisk krop i hvile, der for hvert minut bruger 250 mL ilt og har indtaget en normal blandet vestlig kost, sådan at den respiratoriske kvotient er 0,8; så betyder det, at der hvert minut skal udskilles 200 mL CO2 med respirationen eller som (ideal)gas 7,9 mmol.
Haldane-effekten og carbamino-forbindelser
Det giver intuitivt mening, når man kigger på ligevægten mellem CO2 og vand i blodet, at der må findes både bikarbonat og CO2 i blodet. Den tredje større pulje af CO2 transporteres bundet til amin-grupper på hæmoglobin. Så hvad er det?
Man husker måske Bohr-effekten (ikke denne Bohr, heller ikke denne Bohr, eller denne Bohr, eller denne Bohr, denne Bohr, denne Bohr, eller denne Bohr eller denne Bohr, ej heller denne Bohr, men denne Bohr): Hæmoglobin transporterer oxygen og har højt affinitet for oxygen. I CO2-rige-miljøer er det til gengæld omvendt: CO2 stabiliserer T-formen af hæm-molekylet, sådan at affinitet for oxygen falder. Hæmoglobin dumper simpelthen oxygen, hvor forbruget af oxygen (målt som produktion af CO2) er højt.
Den stakkels, men ekstremt produktive og tilsyneladende meget kløgtige fysiolog John Scott Haldane fandt den modsatte effekt. Desværre for Haldane er Bohr så kendt, at man ikke kan sige “the Haldane effect” uden samtidig at pippe “a reverse Bohr effect”. Det modsatte er ikke tilfældet: Bohr-effekten er en effekt i egen ret.
Så Haldane-effekten er det fund, at i iltrige miljøer mister hæmoglobin affinitet for CO2. Normalt bindes CO2 til amin-grupper på hæm-molekylet i en ikke-katalyseret ligevægt, der kan skrives sådan her:
Hb-NH2 + CO2 ↔ Hb-NH-COO– + H+
Effekten er til at tage at føle på: Deoxygeneret hæm har en godt 3 gange højere affinitet for CO2 end oxygeneret hæm har.
Haldane-effekten har klinisk konsekvens, da den driver en stor del, omtrent 30 %, af den patologiske CO2-ophobning hos patienter med KOL, der modtager ilttilskud. Den ekstra ilt øger oxygeneringen af blodet med den (forudsigelige) konsekvens, at hæmoglobin dumper CO2. Figuren herunder er CO2’s dissociationskurve og beskriver sammenhængen mellem CO2-trykket og CO2-indholdet i blodet. Haldane-effekten er det fund, at ændringer i hæmoglobins iltmætning forskyder dissociationskurven enten opad og til venstre (ved lav iltmætning) eller nedad og til højre (ved høj iltmætning).
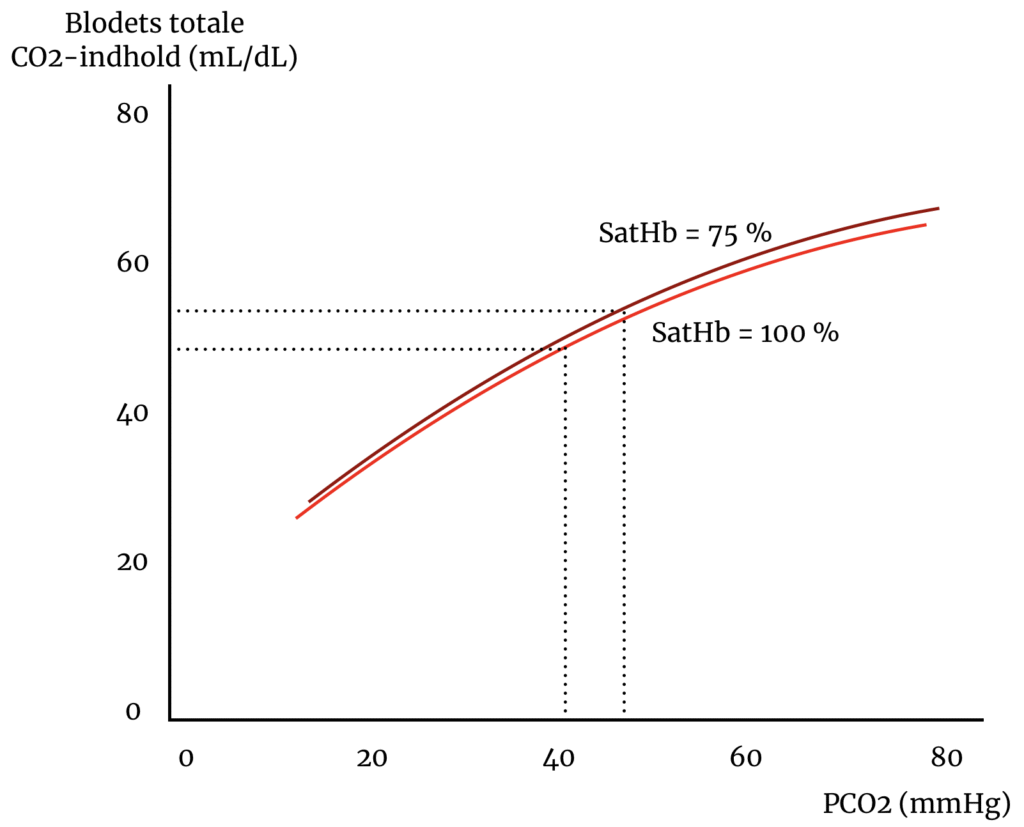
En KOL-patient, der modtager ilt, så iltsaturationen af hæmoglobin bringes fra 75 % til 100 % vil opleve en forskydning af kurven fra den mørkerøde til den lysere røde. CO2-dumpes fra hæmoglobin; en del vil omdannes til bikarbonat og en fraktion til frit, opløst CO2, hvorfor det samme totale CO2-indhold, på grund af Haldane-effekten, vil være ensbetydende med en stigning i CO2-trykket.
Forskelle mellem CO2 i arterielt og i venøst blod
Det fremgår allerede af tabellen ovenfor, at CO2 findes på tre forskellige former i kroppen. Det er ikke helt rigtigt. Der findes mikromolære koncentrationer af kulsyre og karbonat, som jeg i det følgende tillader mig at ignorere. Der er vigtige (og klinisk relevante) forskelle på arterielt blod og venøst blod. Der er også forskelle på, om man befinder sig i plasma, eller om man befinder sig i erytrocytterne, hvor hæmoglobin findes i rigt mål og derfor kan være en kilde til carbamino-forbindelser, og hvor kulsyreanhydrase (= carboanhydrase) også findes i rigt mål.
For den mere visuelt orienterede læser, er her en omsætning af tallene i ovenstående tabel til et fint søjlediagram, der viser de tre hovedfraktioner af CO2 i henholdsvist arterielt og venøst blod.
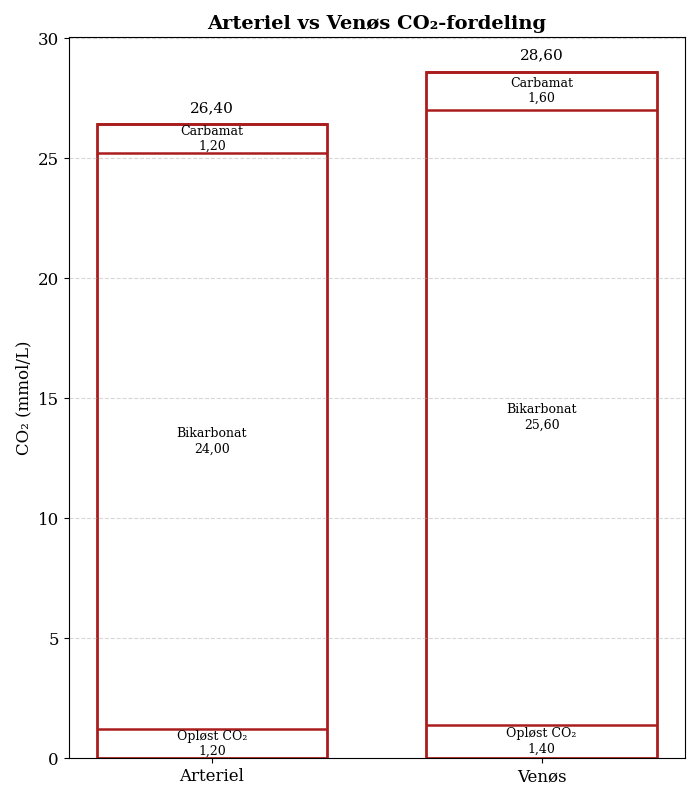
Man bider mærke i, at CO2-indholdet i venøst blod er højere end CO2-indholdet i arterielt blod; omtrent 2,2 mmol/L.
Man bider imidlertid også mærke i, at der ikke sker en proportionel fordeling af den ekstra CO2 til de forskellige fraktioner. Bikarbonat udgør 90 % af den totale CO2 i arterielt blod, men overtager ikke 90 % af den ekstra CO2.
Af de ekstra 2,2 mmol/L bliver, i runde tal, 70 % omdannet til bikarbonat; 20 % til carbamat-forbindelser og 10 % til opløst CO2.
Carboanhydrase katalyserer hydreringen af CO2 (og gør det ekstremt hurtigt)
Helt centralt i transporten af CO2 i blodet er carboanhydrase. Givet den ufattelige interesse alle mulige andre enzymer i kroppen får, er det nok ikke helt forkert at kalde carboanhydrase en unsung hero of metabolism. Der er tale om en gruppe af isoforme enzymer, der alle katalyserer den samme proces som illustreret ved nedenstående figur:
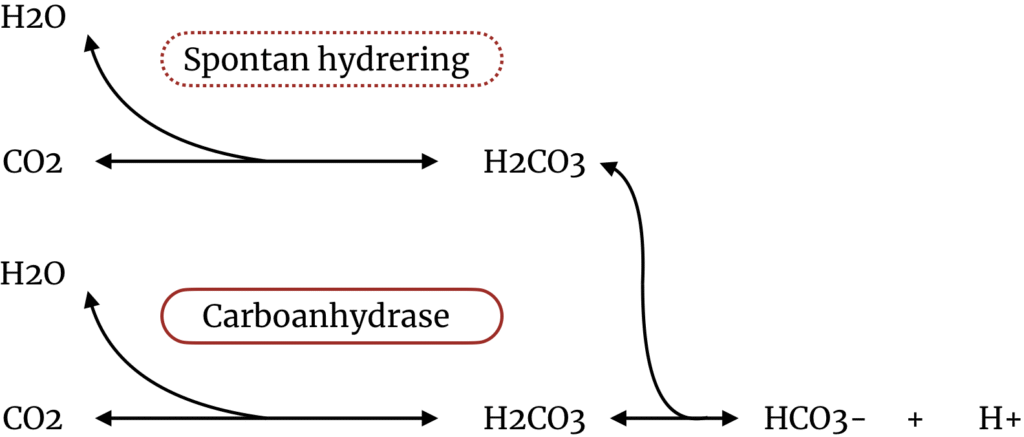
Er man særligt interesseret, kunne man orientere sig i Maren (1967), som beskæftiger sig i store længder med at opsummere, hvad der på daværende tidspunkt var kendt om enzymet, dets fysiologi og inhibering, og som i modsætning til nyere oversigtsartikler er gratis. Jeg har ikke læst det hele, men har formået, med min sparsomme biokemiske baggrund, at hive følgende påstande ud af artiklen. Jeg gengiver dem her i håb om deres rigtighed, og i håb om, at jeg ikke har misforstået for meget. Grundbudskabet, som med sikkerhed er rigtigt er, at den spontane forbindelse mellem CO2 og H2O er relativt langsom, og at carboanhydrase massivt speeder processen op.
Den normale proces er så langsom, at man ikke kan nå ækvillibrium i et normalt respirerende pattedyr. Det betyder kort sagt, at den nødvendige CO2 ikke kan nå at omdannes til bikarbonat uden at CO2-trykket stiger til faretruende højder. Til eksempel: En hvilende hund udleder 50 mL eller 2200 mikromol CO2 i minuttet, hvoraf har cirkuleret som bikarbonat. Den ikke-katalyserede omdannelse af bikarbonat til CO2 er blot 45 mikromol i minuttet. Carboanhydrase øger hastigheden 13.000 (tretten tusinde) gange, sådan at der kan omdannes 600.000 mikromol i minuttet. (Hos Yartsev møder man tallet 25.000 gange). Fra et funktionelt perspektiv må det betyde, at ligevægt skulle man tilføje fx 135 mikromol CO2 (2200 mikromol per minut fordelt på 16 ventilationer i minuttet), ville indtræde på under en tusindedel af et sekund. Respirationsfysiologien opererer jo som bekendt typisk med minutter, timer og dage, så fra et fysiologisk perspektiv betyder det, at ligevægt indtræder momentant.
At der er en betragtelig residualkapacitet understreges også af, at man kan spise carboanhydrase-hæmmere (azetazolamid/diamox) uden at dø af det. Skulle man være så uheldig at inhibere samtlige carboanhydraser i kroppen, ville respirationen skulle bero på den spontane reaktion. Man forestiller sig usundt høje niveauer af frit kuldioxid og svær metabolisk acidose, fordi blodets bufferkapacitet falder.
CO2-dissociationskurven
En længere skrivelse om CO2-dissociationskurven kan findes her.
Kilder
Geers C, Gros G. Carbon dioxide transport and carbonic anhydrase in blood and muscle. Physiol Rev. 2000 Apr;80(2):681-715. doi: 10.1152/physrev.2000.80.2.681. PMID: 10747205. (Graaaaatis!)
Maren TH. Carbonic anhydrase: chemistry, physiology, and inhibition. Physiol Rev. 1967 Oct;47(4):595-781. doi: 10.1152/physrev.1967.47.4.595. PMID: 4964060. (Gratis, lang og ældre)
West JB. Ch 6 Gas Transport by the Blood. I Respiratory Physiology – The Essentials 9th Ed. Wolter Kluwer, 2012, pp.: 77-94.
Boron WF. Ch 29 Transport of Oxygen and Carbon Dioxide in the Blood. I Boron & Boulpaeps Medical Physiology, 3rd Ed. Elsevier, 2016.
Yartsev A (2024). Transport of carbon dioxide in the blood. På DerangedPhysiology.com. Tilgået 24/10-2025.